Péptida nyaéta kelas sanyawa anu dibentuk ku sambungan sababaraha asam amino ngaliwatan beungkeut péptida.Aranjeunna ubiquitous dina organisme hirup.Nepi ka ayeuna, puluhan rébu péptida geus kapanggih dina organisme hirup.Péptida maénkeun peran anu penting dina ngatur kagiatan fungsional rupa-rupa sistem, organ, jaringan sareng sél sareng dina kagiatan kahirupan, sareng sering dianggo dina analisa fungsional, panalungtikan antibodi, pamekaran ubar sareng widang anu sanés.Kalayan ngembangkeun biotéhnologi sareng téknologi sintésis péptida, beuki seueur ubar péptida parantos dikembangkeun sareng diterapkeun di klinik.
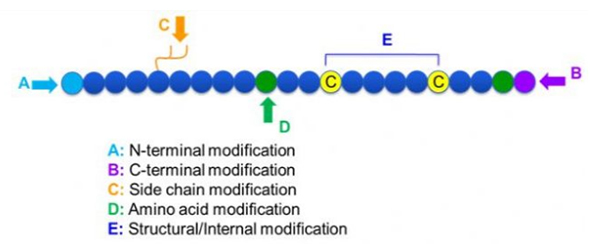
Aya rupa-rupa modifikasi péptida, nu bisa saukur dibagi kana modifikasi pos jeung modifikasi prosés (ngagunakeun modifikasi asam amino diturunkeun), sarta modifikasi N-terminal, modifikasi C-terminal, modifikasi ranté samping, modifikasi asam amino, modifikasi rorongkong, jsb, gumantung kana situs modifikasi (Gambar 1).Salaku sarana penting pikeun ngarobah struktur ranté utama atawa grup ranté samping ranté péptida, modifikasi péptida éféktif bisa ngarobah sipat fisik jeung kimia sanyawa péptida, ngaronjatkeun kaleyuran cai, manjangkeun waktu Peta di vivo, ngarobah distribusi biologis maranéhanana, ngaleungitkeun immunogenicity. , ngurangan efek samping toksik, jsb Dina makalah ieu, sababaraha strategi modifikasi péptida utama jeung ciri maranéhanana diwanohkeun.
1. Siklisasi
Péptida siklik ngagaduhan seueur aplikasi dina biomedis, sareng seueur péptida alami kalayan kagiatan biologis nyaéta péptida siklik.Kusabab péptida siklik condong leuwih kaku ti péptida linier, aranjeunna pisan tahan ka sistem pencernaan, bisa salamet dina saluran pencernaan, sarta némbongkeun afinitas kuat pikeun reséptor target.Siklisasi mangrupikeun cara anu paling langsung pikeun nyintésis péptida siklik, khususna pikeun péptida anu kerangka strukturna ageung.Nurutkeun kana mode cyclization, éta bisa dibagi kana tipe ranté-sisi ranté samping, terminal - tipe ranté samping, terminal - tipe terminal (tungtung ka tungtung tipe).
(1) sidechain-to-sidechain
Jinis anu paling umum tina siklisasi ranté sisi ka ranté sisi nyaéta ngahubungkeun disulfida antara résidu sistein.Siklisasi ieu diwanohkeun ku sapasang résidu sistein anu diproteksi tuluy dioksidasi pikeun ngabentuk beungkeut disulfida.Sintésis polisiklik tiasa dihontal ku cara ngaleungitkeun gugus panyalindungan sulfhydryl sacara selektif.Cyclization bisa dipigawé boh dina pangleyur pos-disosiasi atawa dina résin pre-disosiasi.Siklisasi dina résin bisa jadi kurang éféktif batan siklisasi pangleyur sabab péptida dina résin henteu gampang ngabentuk konformasi siklik.Jinis séjén tina siklisasi ranté samping - ranté samping nyaéta kabentukna struktur amida antara asam aspartat atawa résidu asam glutamat jeung asam amino basa, nu merlukeun gugus panyalindungan ranté samping kudu bisa sacara selektif dikaluarkeun tina polipéptida. dina résin atawa sanggeus disosiasi.Jinis katilu ranté samping - siklisasi ranté samping nyaéta formasi éter diphenyl ku tirosin atanapi p-hydroxyphenylglycine.Siklisasi jinis ieu dina produk alami ngan ukur aya dina produk mikroba, sareng produk siklisasi sering gaduh nilai ubar poténsial.Nyiapkeun sanyawa ieu merlukeun kaayaan réaksi unik, jadi maranéhna teu mindeng dipaké dina sintésis péptida konvensional.
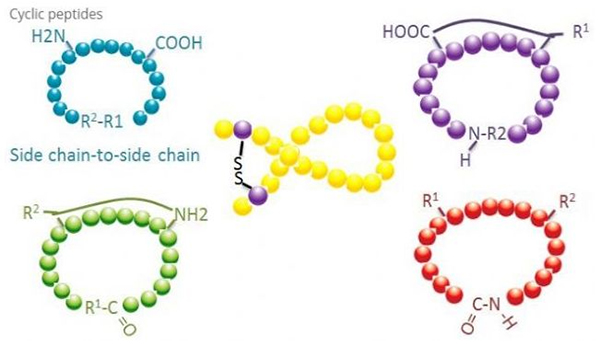
(2) terminal-to-sidechain
Siklisasi ranté sisi terminal biasana ngalibatkeun terminal C sareng gugus amino tina lisin atanapi ornithine ranté samping, atanapi N-terminal sareng asam aspartat atanapi ranté samping asam glutamat.Siklisasi polipéptida séjén dijieun ku cara ngabentuk beungkeut éter antara terminal C jeung sérin atawa treonin ranté samping.
(3) Terminal atawa tipe sirah-to-buntut
Polipéptida ranté tiasa didaur dina pangleyur atanapi dipasang dina résin ku siklik ranté samping.Konsentrasi péptida anu rendah kedah dianggo dina sentralisasi pelarut pikeun nyegah oligomerisasi péptida.Ngahasilkeun polipéptida ring sintétik sirah-ka-buntut gumantung kana runtuyan polipéptida ranté.Ku alatan éta, saméméh Nyiapkeun péptida siklik dina skala badag, hiji perpustakaan mungkin ranté péptida timah mimiti kudu dijieun, dituturkeun ku cyclization pikeun manggihan runtuyan jeung hasil pangalusna.
2. N-métilasi
N-metilasi asalna dina péptida alam sarta diwanohkeun kana sintésis péptida pikeun nyegah kabentukna beungkeut hidrogén, sahingga péptida leuwih tahan ka biodegradasi jeung clearance.Sintésis péptida ngagunakeun turunan asam amino N-methylated nyaéta métode anu paling penting.Salaku tambahan, réaksi Mitsunobu tina N-(2-nitrobenzene sulfonyl chloride) antara polipéptida-résin sareng métanol ogé tiasa dianggo.Métode ieu dipaké pikeun nyiapkeun perpustakaan péptida siklik anu ngandung asam amino N-methylated.
3. Fosforilasi
Fosforilasi mangrupikeun salah sahiji modifikasi pasca-translasi anu paling umum di alam.Dina sél manusa, leuwih ti 30% protéin difosforilasi.Fosforilasi, utamana fosforilasi malik, muterkeun hiji peran penting dina ngadalikeun loba prosés sélular, kayaning transduksi sinyal, éksprési gén, siklus sél jeung régulasi sitoskeleton, sarta apoptosis.
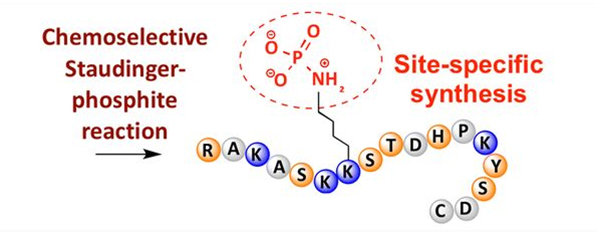
Fosforilasi bisa dititénan dina rupa-rupa résidu asam amino, tapi target fosforilasi nu paling umum nyaéta sérin, treonin, jeung résidu tirosin.Turunan fosfotirosin, fosfotreonin, sareng fosfoserin tiasa dilebetkeun kana péptida salami sintésis atanapi dibentuk saatos sintésis péptida.Fosforilasi selektif bisa dihontal ngagunakeun résidu sérin, threonine, jeung tirosin anu sacara selektif nyabut gugus pelindung.Sababaraha réagen fosforilasi ogé tiasa ngenalkeun gugus asam fosfat kana polipéptida ku modifikasi pasca.Dina taun anyar, fosforilasi situs-spésifik lisin geus kahontal maké réaksi Staudinger-phosphite selektif kimiawi (Gambar 3).
4. Myristoylation na palmitoylation
Asilasi terminal N jeung asam lemak ngamungkinkeun péptida atawa protéin ngabeungkeut mémbran sél.Runtuyan myridamoylated dina N-terminal ngamungkinkeun kinase protéin kulawarga Src jeung protéin Gaq transcriptase sabalikna bisa jadi sasaran pikeun ngabeungkeut mémbran sél.Asam myristic ieu numbu ka N-terminal résin-polipéptida maké réaksi gandeng baku, sarta hasilna lipopeptide bisa disosiasi dina kaayaan baku sarta dimurnikeun ku RP-HPLC.
5. Glikosilasi
Glikopéptida sapertos vankomisin sareng teikolanin mangrupikeun antibiotik anu penting pikeun pengobatan inféksi baktéri anu tahan obat, sareng glikopéptida sanés sering dianggo pikeun merangsang sistem imun.Salaku tambahan, sabab seueur antigén mikroba anu glikosilasi, penting pisan pikeun diajar glikopéptida pikeun ningkatkeun pangaruh terapi inféksi.Di sisi séjén, geus kapanggih yén protéin dina mémbran sél sél tumor némbongkeun glikosilasi abnormal, nu ngajadikeun glikopéptida maénkeun peran penting dina kanker sarta panalungtikan pertahanan imun tumor.Glikopéptida disusun ku métode Fmoc/t-Bu.Résidu glikosilasi, sapertos treonin sareng serin, sering diwanohkeun kana polipéptida ku fMOCs éster pentafluorophenol anu diaktipkeun pikeun ngajaga asam amino glikosilasi.
6. Isoprena
Isopentadienylation lumangsung dina résidu sistein dina ranté samping deukeut C-terminal.Protéin isoprene tiasa ningkatkeun afinitas mémbran sél sareng ngabentuk interaksi protéin-protéin.Protéin anu diisopentadienasi kalebet tirosin fosfatase, GTase leutik, molekul cochaperone, lamina nuklir, sareng protéin pengikat centromeric.Polipéptida isoprena tiasa disiapkeun nganggo isoprena dina résin atanapi ku ngawanohkeun turunan sistein.
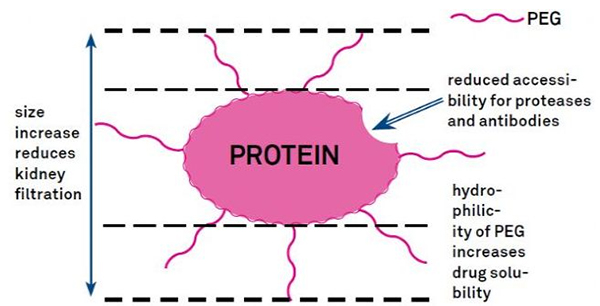
7. Poliétilén glikol (PEG) modifikasi
Modifikasi PEG tiasa dianggo pikeun ningkatkeun stabilitas hidrolitik protéin, biodistribusi sareng kaleyuran péptida.Perkenalan ranté PEG kana péptida tiasa ningkatkeun sipat farmakologisna sareng ogé ngahambat hidrolisis péptida ku énzim proteolitik.PEG péptida ngaliwatan bagian melintang kapiler glomerulus leuwih gampang batan péptida biasa, greatly ngurangan bersihan ginjal.Kusabab satengah-umur aktif péptida PEG dina vivo, tingkat perlakuan normal tiasa dipertahankeun kalayan dosis anu langkung handap sareng obat péptida anu kirang sering.Sanajan kitu, modifikasi PEG ogé boga épék négatip.Jumlah PEG anu ageung nyegah énzim ngahina péptida sareng ogé ngirangan beungkeutan péptida kana reséptor target.Tapi afinitas low péptida PEG biasana diimbangi ku satengah hirup farmakokinetikna anu langkung panjang, sareng ku ayana dina awak langkung lami, péptida PEG gaduh kamungkinan anu langkung ageung diserep kana jaringan target.Ku alatan éta, spésifikasi polimér PEG kudu dioptimalkeun pikeun hasil optimal.Di sisi séjén, péptida PEG akumulasi dina ati alatan ngurangan clearance ginjal, hasilna sindrom makromolekul.Ku alatan éta, modifikasi PEG kudu dirancang leuwih taliti lamun péptida dipaké pikeun nguji ubar.
Grup modifikasi umum tina modifiers PEG bisa kasarna diringkeskeun kieu: Amino (-amine) -NH2, aminomethyl-Ch2-NH2, hydroxy-OH, carboxy-Cooh, sulfhydryl (-Thiol) -SH, Maleimide -MAL, succinimide karbonat - SC, succinimide acetate -SCM, succinimide propionate -SPA, n-hydroxysuccinimide -NHS, Acrylate-ch2ch2cooh, aldehyde -CHO (sapertos propional-ald, butyrALD), basa akrilik (-acrylate-acrl), azido-azide, biotinyl - Biotin, Fluorescein, glutaryl -GA, Acrylate Hydrazide, alkuna-alkuna, p-toluenesulfonate -OTs, succinimide succinate -SS, jsb Turunan PEG kalawan asam karboksilat bisa gandeng kana n-terminal amina atawa ranté samping lisin.Amino-diaktipkeun PEG bisa gandeng kana asam aspartat atawa asam glutamat ranté samping.Mal-diaktipkeun PEG bisa conjugated kana mercaptan tina ranté samping sistein pinuh deprotected [11].Modifier PEG biasana digolongkeun kieu (catetan: mPEG nyaéta methoxy-PEG, CH3O-(CH2CH2O)n-CH2CH2-OH):
(1) ranté lempeng PEG modifier
mPEG-SC, mPEG-SCM, mPEG-SPA, mPEG-OTs, mPEG-SH, mPEG-ALD, mPEG-butyrALD, mPEG-SS
(2) modifier PEG bifunctional
HCOO-PEG-COOH, NH2-PEG-NH2, OH-PEG-COOH, OH-PEG-NH2, HCl·NH2-PEG-COOH, MAL-PEG-NHS
(3) branching PEG modifier
(mPEG)2-NHS, (mPEG)2-ALD, (mPEG)2-NH2, (mPEG)2-MAL
8. Biotinisasi
Biotin tiasa ngabeungkeut pisan sareng avidin atanapi streptavidin, sareng kakuatan beungkeutna caket sareng beungkeut kovalén.Péptida anu dilabélan biotin biasana dianggo dina immunoassay, histocytochemistry, sareng sitometri aliran dumasar-fluoresensi.Antibodi antibiotin anu dilabélan ogé tiasa dianggo pikeun ngabeungkeut péptida anu dibiotinilasi.labél biotin mindeng napel ranté samping lisin atawa terminal N.Asam 6-aminocaproic sering dianggo salaku beungkeut antara péptida sareng biotin.Beungkeutna fleksibel dina ngabeungkeut substrat sareng ngabeungkeut langkung saé upami aya halangan sterik.
9. labél fluoresensi
Labél fluoresensi tiasa dianggo pikeun ngalacak polipéptida dina sél hirup sareng ngulik énzim sareng mékanisme tindakan.Tryptophan (Trp) nyaéta fluoresensi, ku kituna tiasa dianggo pikeun panyiri intrinsik.Spéktrum émisi triptofan gumantung kana lingkungan periferal sareng turun kalayan ngirangan polaritas pelarut, sipat anu kapaké pikeun ngadeteksi struktur péptida sareng beungkeutan reséptor.Fluoresensi triptofan tiasa dipadamkeun ku asam aspartat anu diprotonasi sareng asam glutamat, anu tiasa ngabatesan panggunaanana.Gugus klorida Dansyl (Dansyl) kacida fluoresensi lamun kabeungkeut kana gugus amino sarta mindeng dipaké salaku labél fluoresensi pikeun asam amino atawa protéin.
Konversi énergi résonansi fluoresensi (FRET) mangpaat pikeun studi énzim.Nalika FRET diterapkeun, polipéptida substrat biasana ngandung gugus panyiri fluoresensi sareng gugus panyiri fluoresensi.Grup fluoresensi anu dilabélan dipaénkeun ku pamaen ngaliwatan transfer énergi non-foton.Nalika péptida dipisahkeun tina énzim anu dimaksud, grup panyiri ngaluarkeun fluoresensi.
10. Polipéptida kandang
Péptida kandang gaduh gugus pelindung anu tiasa dicabut sacara optik anu ngalindungan péptida tina ngariung kana reséptor.Nalika kakeunaan radiasi UV, péptida diaktipkeun, mulangkeun pangirutna ka reséptor.Kusabab aktivasina optik ieu bisa dikawasa nurutkeun waktu, amplitudo, atawa lokasi, péptida kandang bisa dipaké pikeun nalungtik réaksi lumangsung dina sél.Grup pelindung anu paling sering dianggo pikeun polipéptida kandang nyaéta gugus 2-nitrobenzyl sareng turunanna, anu tiasa diwanohkeun dina sintésis péptida ngalangkungan turunan asam amino pelindung.Turunan asam amino anu geus dimekarkeun nyaéta lisin, sistein, sérin, jeung tirosin.Turunan aspartat sareng glutamat, kumaha oge, henteu biasa dianggo kusabab karentanan kana siklisasi nalika sintésis péptida sareng disosiasi.
11. Péptida Polyantigenic (MAP)
Péptida pondok biasana teu imun jeung kudu gandeng jeung protéin pamawa pikeun ngahasilkeun antibodi.Péptida Polyantigenic (MAP) diwangun ku sababaraha péptida idéntik disambungkeun ka inti lisin, nu husus bisa nganyatakeun immunogens potency tinggi tur bisa dipaké pikeun nyiapkeun couplets protéin péptida-carrier.Polipéptida MAP tiasa disintésis ku sintésis fase padet dina résin MAP.Tapi, kopling anu teu lengkep nyababkeun ranté péptida leungit atanapi dipotong dina sababaraha cabang sahingga henteu nunjukkeun sipat polipéptida MAP asli.Alternatipna, péptida tiasa disiapkeun sareng dimurnikeun sacara misah teras gandeng kana MAP.Runtuyan péptida napel inti péptida geus well-diartikeun sarta gampang dicirikeun ku spéktrometri massa.
kacindekan
Modifikasi péptida mangrupikeun cara anu penting pikeun ngarancang péptida.Péptida anu dirobih sacara kimia henteu ngan ukur tiasa ngajaga kagiatan biologis anu luhur, tapi ogé sacara efektif ngahindarkeun kakurangan immunogenicity sareng karacunan.Dina waktos anu sami, modifikasi kimiawi tiasa masihan péptida ku sababaraha sipat énggal anu saé.Dina taun-taun ayeuna, metode aktivasina CH pikeun modifikasi pasca polipéptida parantos gancang dikembangkeun, sareng seueur hasil anu penting parantos kahontal.
waktos pos: Mar-20-2023